- Теломераза
-
Теломераза — фермент, добавляющий особые повторяющиеся последовательности ДНК (TTAGGG у позвоночных) к 3'-концу цепи ДНК на участках теломер, которые располагаются на концах хромосом в эукариотических клетках. Теломеры содержат уплотненную ДНК и стабилизируют хромосомы. При каждом делении клетки теломерные участки укорачиваются. Существование механизма, компенсирующего укорочение теломер (теломеразы), было предсказано в 1973 году А. М. Оловниковым[1].
Теломераза является обратной транскриптазой, причем с ней связана особая молекула РНК, которая используется в качестве матрицы для обратной транскрипции во время удлинения теломер.
Теломераза была обнаружена Кэрол Грейдер в 1984 году[2].
За открытие защитных механизмов хромосом от концевой недорепликации с помощью теломер и теломеразы в 2009 году присуждена Нобелевская премия по физиологии и медицине австралийке, работающей в США, Элизабет Блэкбёрн, американке Кэрол Грейдер и её соотечественнику Джеку Шостаку.[3]
Содержание
Общие сведения
В результате деятельности теломеразы длина теломерных участков хромосом клетки увеличивается или сохраняется на постоянном уровне, компенсируя таким образом концевую недорепликацию и позволяя клетке делиться неограниченно долго. В ходе исследования этого фермента (состоящего, как описано ниже, из РНК-компонента и белкового компонента) выяснилось, что РНК-компонент экспрессируется на постоянном уровне практически во всех клетках, и для индуцирования теломеразной активности необходима экспрессия белкового компонента, названного поэтому каталитическим компонентом теломеразы. Искусственно индуцированная экспрессия гена каталитического компонента теломеразы (путем введения гена при помощи методов генной инженерии) делает клеточную культуру иммортальной (бессмертной), то есть способной делиться неограниченно долго, отменяя тем самым для культуры предел Хейфлика. Теломераза экспрессируется в стволовых, половых и некоторых других типах клеток организма, которым необходимо постоянно делиться для функционирования определённых тканей (например, клетки эпителия кишечника). Обычные соматические клетки организма лишены теломеразной активности. Клетки 85 % раковых опухолей обладают теломеразной активностью, поэтому считается, что активация теломеразы является одним из событий на пути клетки к злокачественному перерождению.
Структура
Строение человеческой теломеразы было выяснено Скоттом Коэном и его исследовательской группой в австралийском Исследовательском институте детской медицины. Теломераза состоит из теломеразной обратной транскриптазы (TERT), теломеразной РНК (hTR или TERC), и дискерина (по две молекулы каждого из этих веществ) [4]. Две субъединицы фермента кодируются двумя различными генами. Кодирующий регион для TERT имеет длину 3396 пар, TERT таким образом содержит 1131 аминокислоту. Этот белок сворачивается и захватывает TERC (имеющую длину 451 нуклеотид), которая не транслируется, оставаясь РНК. TERT имеет форму рукавицы, что позволяет ему закрепляться на хромосоме и добавлять в нее одноцепочечные теломерные участки.
TERT — это обратная транскриптаза, то есть фермент, создающий одноцепочечную ДНК на основе шаблонной одноцепочечной РНК. Такие ферменты (а именно — образцы, полученные из вирусов) используются исследователями в лабораторном процессе полимеразной цепной реакции обратной транскрипции (RT-PCR), при помощи которого создаются копии ДНК на основе заданного РНК-шаблона. TERT же удерживает свой собственный шаблон — TERC.
Профессор Скордалакес и его коллеги первыми выявили полный набор и структуру критически важных белков в молекуле теломеразы. [5]
Назначение
Используя TERC, TERT добавляет повторяющуюся последовательность из шести нуклеотидов 5'-TTAGGG к 3'-нити хромосом. Указанная последовательность используется у позвоночных; у других классов организмов последовательности отличаются. Эти повторяющиеся последовательности TTAGGG вместе со своими парными белками, и называются теломерами.
Шаблонный участок TERC содержит последовательность 3'-AAUCCC-5'. Теломераза связывает первые несколько нуклеотидов шаблона с последней теломерной последовательностью на хромосоме, добавляет новый повторяющийся участок (5'-TTAGGG-3'), отделяется, связывает новый 3'-конец теломеры с шаблоном и повторяет весь процесс заново.
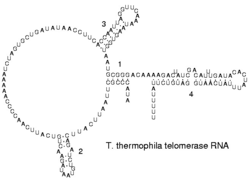 Вторичная структура РНК-компонента теломеразы инфузории Tetrahymena termophila
Вторичная структура РНК-компонента теломеразы инфузории Tetrahymena termophila
Объяснение необходимости удлинения теломер приведено в основной статье о теломерах.
Клинические представления
Старение
Теломеразу считают ключом к клеточному бессмертию, «источником юности». Этот фермент позволяет клеткам быстро размножаться без старения. Стволовые клетки эмбрионов, например, экспрессируют теломеразу, которая позволяет им непрерывно делиться, формируя ткани и органы. У взрослых организмов теломераза экспрессируется в клетках, которые должны часто делиться, однако большинство соматических клеток ее не производят.
Один из наиболее очевидных признаков старости — сниженная активность клеток кожи. Предполагается, что лечение теломеразой сможет помочь избавиться по крайней мере от этих проблем.
Выяснилось что β-катенин, избежавший деградации благодаря Wnt сигнала, активирует синтез ферментативной субъединицы теломеразы (TERT) в стволовых и раковых клетках. В этом ему помогает один из транскрипционных факторов плюрипотенции — Klf4, направляющий его на промотор гена Tert[6] . Компании Revive Skincare и TaSciences.com выпускают косметические продукты на основе теломераз. По заявлениям Revive, производство одного грамма теломеразы обходится компании в 4 миллиона долларов США. Активной теломеразы в косметических препаратах нет и быть не может, поскольку содержащаяся в составе теломеразы РНК очень быстро разрезается рибонуклеазой А, которая очень распространена в природе. Кроме того и сам фермент не очень стабилен. Активную теломеразу в лабораториях выделяют в стерильных условиях и хранят в замороженном виде (желательно в низкотемпературном холодильнике).
Возникновение многих признаков раннего старения приписывают сокращению теломер[7]. Это, например, прогерия, Атаксия-телангиэктазия, синдром Блума, анемия Фанкони, синдром Наймегена и другие телангиэктазивные нарушения. Гены, мутация которых приводит к этим заболеваниям, задействованы в восстановлении ДНК. Их участие в управлении длиной теломер активно исследуется. Поскольку на настоящий момент неизвестно, насколько разрушение теломер влияет на процесс старения, основные исследования направлены на процессы сохранения целостности ДНК и в особенности ее теломерных участков. Майкл Фоссел в одном из интервью предположил, что лечение теломеразой может использоваться не только для борьбы с раком, но даже для борьбы со старением человеческого организма и таким образом для увеличения продолжительности жизни. Он считает, что уже в ближайшем десятилетии будут проведены первые испытания теломеразных методов увеличения продолжительности жизни человека. Это весьма обнадеживает, так как примерно в это же время произойдет массовый уход на пенсию людей поколения беби-бума в США и западной Европе.
Рак
Когда клетки в культуре приближаются к пределу Хейфлика, старение может быть замедлено деактивацией генов, которые кодируют белки, подавляющие образование опухолей. Это, в частности, белок, называемый p53, и белок ретинобластомы (pRb). Измененные таким образом клетки рано или поздно достигают состояния, называемого «кризисом», когда большая часть клеточной культуры умирает. Иногда, однако, клетка не перестает делиться при достижении кризиса. Обычно в это время теломеры полностью разрушены и состояние хромосомы ухудшается с каждым делением.
Оголенные концы хромосом распознаются как разрывы обеих цепей ДНК. Повреждения такого рода обычно устраняются путем соединения разорванных концов. Однако в данном случае соединенными могут оказаться концы разных хромосом, так как они более не защищены теломерами. Это временно позволяет решить проблему отсутствия теломер, однако во время анафазы клеточного деления сцепленные хромосомы разрываются на части случайным образом, что приводит к большому количеству мутаций и хромосомных аномалий.
По мере продолжения этого процесса геном клетки повреждается все больше. Наконец, наступает момент, когда либо объем поврежденного генетического материала становится достаточным для гибели клетки (путем запрограммированной клеточной смерти — апоптоза), либо происходит дополнительная мутация, активирующая теломеразу.
После активации теломеразы некоторые виды клеток становятся бессмертными: их хромосомы не становятся менее стабильными вне зависимости от числа клеточных делений, и процесс клеточной смерти не запускается. Многие раковые клетки считаются бессмертными, поскольку активность генов теломеразы в них позволяет им делиться практически бесконечно, что и является причиной образования опухолей.
Хороший пример бессмертия раковых клеток — это клетки HeLa, изначально полученные из опухоли шейки матки Генриетты Лекс (Henrietta Lacks, отсюда название культуры HeLa) в 1951 г. Эта культура по сей день используется в лабораторных исследованиях. Клетки Hela в самом деле бессмертны: по оценкам ежедневно производится несколько тонн этих клеток, причем все они являются потомками нескольких клеток, извлеченных из опухоли Г. Лекс.
Несмотря на то, что описанный метод моделирования рака в культуре клеток эффективен и используется уже много лет, он весьма неточен. Какое именно воздействие в рамках этого метода вызывает рождение клеток, образующих опухоли, изначально было неясно. Однако впоследствии удалось ответить на этот вопрос. В модельных клетках вызывались различные мутации, которые встречаются в разных видах человеческих раковых клеток. Это позволило выявить несколько сочетаний мутаций, которые являются достаточными для образования опухолевых клеток из различных их видов.
Собственно сочетания мутаций различается в зависимости от вида клеток. Однако большинство из них содержат следующие изменения:
- активацию теломеразы;
- нарушение цикла белка p53;
- активацию прото-онкогенов Ras, Myc или других;
- нарушение формирования фосфатазы белка PP2A.
Эти изменения приводят к отключению механизма гибели клетки в результате разрушения хромосом или апоптоза. Кроме того клетка начинает непрерывно делиться.
Эта модель рака, созданная в клеточной культуре, проливает свет на роль теломеразы в формирование опухолей у человека. Активация теломеразы наблюдается в 90 % всех опухолей. Это позволяет заключить, что предоставляемое ею бессмертие является ключевым фактором в развитии рака.
Опухоли, в клетках которых TERT не активировался, в основном использовали другой механизм сохранения теломер, называемый ALT (альтернативное удлинение теломер, alternative lengthening of telomeres). Детали этого механизма неизвестны.
Как показано в работе Элизабет Блекбёрн и др., теломераза также участвует в регуляции активности 70 генов, которые участвуют либо подозреваются в участии в образовании и развитии раковых опухолей. Кроме того, она активирует гликолиз, что позволяет раковым клеткам использовать сахара для поддержания заданной скорости роста и деления (эти скорости сравнимы со скоростями роста клеток в зародыше).
Э. Блэкбёрн и ещё двое исследователей получили премию Ласкера в 2006 г. за открытие теломеразы и последующие ее исследования. За те же достижения Э. Блэкбёрн также получила премию Грубера по генетике в 2006 г.
Роль теломеразы в других болезнях человека
Считается, что мутации белка TERT вызывают у пациентов предрасположенность к апластической анемии — нарушению кроветворения в костном мозге[8].
Синдром кошачьего крика (CdCS) — сложное расстройство, включающее в себя утрату дистальной части короткого конца пятой хромосомы. Ген белка TERT располагается в утраченном фрагменте. Предполагается, что потеря этой копии гена является если не причиной, то одним из условий возникновения этой болезни[9].
Врожденный дискератоз (ВД) — болезнь костного мозга, которая может быть вызвана мутацией в TERC-субъединице теломеразы. Эта мутация является причиной болезни лишь в 5 % случаев, однако в этих случаях болезнь наследуется как доминантное аутосомное расстройство. Мутации в гене дискерина (DKC1) обуславливают примерно 35 % всех случаев ВД; эти мутации — рецессивные Х-связанные.
Пациенты с ВД страдают от серьёзного нарушения функционирования костного мозга, которое проявляется в аномальной пигментации кожи, лейкоплакии (белые бляшки на слизистой ротовой полости), дистрофии ногтей, а также другими симптомами. У носителей мутаций TERC или DKC1 также выявлены более короткие теломеры, а также некорректная активность теломеразы in vitro, по сравнению с особями того же возраста, свободными от мутаций.[10]
Обнаружена целая семья[11], у членов которой аутосомальный доминантный ВД проявился одновременно с гетерозиготной мутацией TERT. У этих людей скорость сокращения теломер была выше нормальной. Кроме того, фенотип ВД ухудшался с каждым поколением.
Теломераза как мишень для лекарственных препаратов
С раковыми опухолями очень тяжело бороться, так как иммунная система организма их не опознает как чужеродные. Кроме того, раковые клетки бессмертны и будут продолжать делиться при любых условиях. Поскольку в большинстве видов опухолей бессмертие клеток достигается за счет присутствия в них теломеразы, именно она могла бы быть целью для препаратов против рака. Если какой-либо препарат сможет отключить теломеразу в раковых клетках, процесс сокращения теломер возобновится, по мере деления клеток теломеры разрушатся, возникнут мутации и клетки погибнут.
Экспериментальные препараты, воздействующие на активную теломеразу, тестируются на мышах, и некоторые уже перешли к клиническим испытаниям. Geron Corporation в настоящее время испытывает на людях препараты, использующих ингибирование теломеразы двумя различными способами. Один — это вакцина (GRNVAC1), а другой — липидный препарат (GRN163L). В действительности во многих типах раковых клеток, выращенных в культуре, подавление теломеразы ведет к быстрой гибели популяции клеток. Между тем существуют определенные препятствия к развитию такого рода лекарств, например, существование альтернативного механизма удлинения теломер[12]. Есть свидетельства, что альтернативные методы поддержания длины теломер и хранения ДНК применены в стволовых раковых клетках. Geron Corp., однако, заявляет, что им удалось уничтожить и стволовые раковые клетки при помощи их ингибитора теломеразы GRN163L. Этот ингибитор присоединяется непосредственно к РНК-шаблону теломеразы. Даже единичная мутация шаблона могла бы полностью лишить теломеразу ее способности удлинять теломеры и таким образом лишить клетку возможности бесконечно делиться. В этом случае не может начаться гликолиз, а также не может начаться экспрессия 70 раковых генов Блэкбёрн (Blackburn et al).
Поскольку Блэкбёрн показала, что большинство негативных эффектов теломеразы исчезают при повреждении РНК-шаблона, этот шаблон является довольно эффективной мишенью. Однако если даже некоторые из раковых клеток используют альтернативный метод удлинения теломер, они не погибнут, если РНК-шаблон теломеразы будет заблокирован. По мнению Блэкбёрн, неправильно считать, что только теломераза отвечает за удлинение теломер. Предотвращение в раковых клетках гликолиза, а также предотвращение экспрессии 70 «плохих» генов, вероятно, может убить раковые клетки, не использующие теломеразу.
Примечания
- ↑ Olovnikov AM. A theory of marginotomy. The incomplete copying of template margin in enzymic synthesis of polynucleotides and biological significance of the phenomenon. J Theor Biol. 1973 Sep 14;41(1):181-90
- ↑ Greider, C.W. & Blackburn, E.H. (1985) «Identification of a specific telomere terminal transferase activity in Tetrahymena extracts.» Cell v.43, (2 Pt. 1) pp. 405—413.
- ↑ Сайт Нобелевской премии.
- ↑ Cohen S, Graham M, Lovrecz G, Bache N, Robinson P, Reddel R (2007). «Protein composition of catalytically active human telomerase from immortal cells». Science 315 (5820): 1850-3. PMID 17395830.
- ↑ Ученые докопались до механизма клеточного бессмертия
- ↑ Hoffmeyer K. , A. Raggioli, S. Rudloff, R. Anton, A. Hierholzer, I. Del Valle, K. Hein, R. Vogt, R. Kemler. Wnt/ β-Catenin Signaling Regulates Telomerase in Stem Cells and Cancer Cells. Science, 2012; 336 (6088): 1549—1554 DOI: 10.1126/science.1218370
- ↑ Blasco MA. Telomeres and human disease: ageing, cancer and beyond. Nat Rev Genet 2005 Aug;6(8):611-22. PMID 16136653
- ↑ Файл:Free text.png Yamaguchi H, Calado RT, Ly H, Kajigaya S, Baerlocher GM, Chanock SJ, Lansdorp PM, Young NS. Mutations in TERT, the gene for telomerase reverse transcriptase, in aplastic anemia. N Engl J Med. 2005 Apr 7;352(14):1413-24. PMID 15814878 Free text after registration
- ↑ Файл:Free text.png Zhang A, Zheng C, Hou M, Lindvall C, Li KJ, Erlandsson F, Bjorkholm M, Gruber A, Blennow E, Xu D.Deletion of the telomerase reverse transcriptase gene and haploinsufficiency of telomere maintenance in Cri du chat syndrome. Am J Hum Genet. 2003 Apr;72(4):940-8. Epub 2003 Mar 10. PMID 12629597
- ↑ Marrone A, Walne A, Dokal I. Dyskeratosis congenita: telomerase, telomeres and anticipation. Curr Opin Genet Dev. 2005 Jun;15(3):249-57. PMID 15917199
- ↑ Armanios M, Chen JL, Chang YP, Brodsky RA, Hawkins A, Griffin CA, Eshleman JR, Cohen AR, Chakravarti A, Hamosh A, Greider CW. Haploinsufficiency of telomerase reverse transcriptase leads to anticipation in autosomal dominant dyskeratosis congenita. Proc Natl Acad Sci U S A. 2005 Nov 1;102(44):15960-4. PMID 16247010
- ↑ Bryan TM, Englezou A, Gupta J, Bacchetti S, Reddel RR. Telomere elongation in immortal human cells without detectable telomerase activity. EMBO J. 1995 Sep 1;14(17):4240-8. PMID 7556065
10. Guenther Witzany (2007). Telomeres in Evolution and Development from Biosemiotic Perspective. Nature Precedings: doi:10.1038/npre.2007.932.1
См. также
Внешние ссылки
- The Telomerase Database — A Web-based tool for telomerase research.
- Telomerase.org — A blog-style discussion of telomerase and anti-aging therapies.
- Three-dimensional model of telomerase at Memorial University of Newfoundland
 Категории:
Категории:- Старение
- ДНК
- Ферменты
{kind=link}
Wikimedia Foundation. 2010.