- Археоптерикс
-
Не следует путать с Археоптерис.
† Археоптерикс  Реконструкция археоптерикса,
Реконструкция археоптерикса,
музей Оксфордского университетаНаучная классификация Царство: Животные Тип: Хордовые Подтип: Позвоночные Класс: Птицы Отряд: Archaeopterygiformes Семейство: Archaeopterygidae Род: Археоптерикс Вид: A. lithographica Латинское название Archaeopteryx lithographica Meyer, 1861 Археопте́рикс (лат. Archaeopteryx; греч. ἀρχαῖος — древний, πτέρυξ — крыло) — вымершая птица позднего Юрского периода, величиной с ворону, занимающая по морфологии промежуточное положение между пресмыкающимися и птицами. Самый примитивный и ранний из всех известных представителей птиц[1], обитавший 150—147 млн лет назад. Все находки археоптерикса относятся к окрестностям Зольнхофена на юге Германии.
По строению таза и рёбер, по двояковогнутым позвонкам, челюстям, покрытым зубами, и по длинному хвосту, состоящему из более 20 позвонков, этот вид тесно примыкает к пресмыкающимся. Но некоторые аспекты строения конечностей и черепа типичны для птиц; передние конечности снабжены сильными маховыми перьями, хвостовые позвонки также снабжены попарно сидящими на них перьями; наконец, всё тело археоптерикса покрыто перьями.
Содержание
История открытия
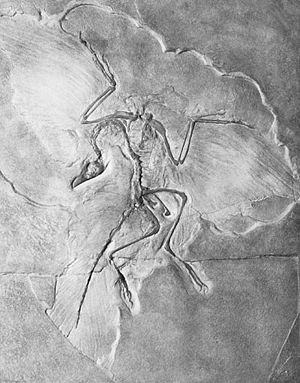 Берлинский экземпляр. Фото 1877 г.
Берлинский экземпляр. Фото 1877 г.
К началу XXI века обнаружено десять скелетных экземпляров археоптерикса и отпечаток пера. Все находки относятся к тонкослоистым известнякам позднеюрского возраста возле Зольнхофена в Баварии.
Список находок, имеющих отношение к археоптериксам:
- Перо.
Обнаружено в 1861 году около Зольнхофена, описано в 1861 году Германом фон Майером. Находится в музее естествознания в Берлине. В основном отождествляется с археоптериксом, поскольку других птиц в Зольнхофенских отложениях не найдено. Является первоначальным голотипом для рода Archaeopteryx и вида A. lithographica.
- Лондонский экземпляр (BMNH 37001, голотип).
Обнаружен в 1861 году около Лангенальтайма. Описан в 1863 году Ричардом Оуэном, который сделал его типовым экземпляром для вида и рода. Хранится в музее естествознания в Лондоне. Голова отсутствует. Реставрация головы была проведена Ричардом Оуэном и позднее подтвердилась после находки берлинского экземпляра в 1877 году.
- Берлинский экземпляр (HMN 1880).
Обнаружен в 1876 или 1877 на Blumenberg, около Айхштета. Якоб Нимейер (Jakob Niemeyer) выменял за корову драгоценные останки у Йохана Дорра (Johann Dörr). Экземпляр описан в 1884 году Вильгельмом Дамесом (Wilhelm Dames). Хранится в музее естествознания в Берлине. Был классифицирован Деймсом как новый вид, A. siemensii в честь Вернера фон Сименса.
- Максбергский экземпляр (S5).
Только туловище. Обнаружен в 1956 или 1958 году около Лангенальтайма, описан в 1959 Геллером (Heller). Детально изучен Джоном Остромом в 1970-ых. В течение некоторого времени демонстрировался в Максбергском музее, после чего был возвращен владельцу. После смерти коллекционера в 1991 году экземпляр обнаружить не удалось. Предполагается, что он был тайно продан владельцем или украден.
- Харлемский экземпляр (TM 6428, также известен как тейлеровский экземпляр).
Обнаружен в 1855 году около Риденбурга, описан Мейером как Pterodactylus crassipes в 1875 году, переклассифицирован Джоном Остромом (John Ostrom) в 1970-м. Находится в музее Тейлера, Харлем (Нидерланды). Самый первый экземпляр, если не учитывать ошибку классификации.
- Айхштетский экземпляр (JM 2257).
Обнаружен в 1951 или 1955 около Workerszell. Описал Петер Велнхофер (Peter Wellnhofer) в 1974. Находится в юрском музее города Айхштет. Самый маленький экземпляр. Имеет сохранившуюся голову, хотя и менее сохранившуюся, чем у термополисского экземпляра.
 Мюнхенский экземпляр
Мюнхенский экземпляр- Мюнхенский экземпляр (S6, ранее известный как Solnhofen-Aktien-Verein).
Обнаружен в 1991 году около Лангенальтайма. Описал Велнхофер в 1993. Хранится в Мюнхенском палеонтологическом музее. Единственный экземпляр с грудиной. Возможно, новый вид, A. bavarica.
- Зольнхофенский экземпляр (BSP 1999).
Обнаружен в 1960-х годах около Айхштета. Описал Велнхофер в 1988. Хранится в Музее бургомистра Мюллера, Зольнхофен. Первоначально был классифицирован коллекционером-любителем как компсогнатус (Compsognathus). Может принадлежать к отдельному роду и виду Wellnhoferia grandis.
- Мюллеровский экземпляр.
Фрагментарный экземпляр, обнаруженный в 1997. Хранится в Мюллеровском музее.
- Неописанный экземпляр.
В 1997 Маузер сообщил об обнаружении в частной коллекции фрагментарного экземпляра [2]. Имя владельца и местонахождение окаменелости не разглашаются. До настоящего момента экземпляр формально не классифицирован, его отнесение к археоптериксам остается предварительным.
 Термополиский экземпляр
Термополиский экземпляр- Термополиский экземпляр (WDC-CSG-100).
Обнаружен в Германии. Долгое время был в частной коллекции, описан в 2005 году в журнале Science [3]. Находится в Wyoming Dinosaur Center, Термополис (штат Вайоминг, США). Имеет наиболее сохранившуюся голову и ступни. Более подробно описан в 2007[4].
Скелет
В данном разделе приводится краткий обзор остеологии археоптерикса: описываются только основные детали анатомии скелета. Строение скелета археоптерикса сравнивается со строением скелета современных птиц и дейнонихозавров — тероподных динозавров, являющихся, по мнению большинства палеонтологов, ближайшими родственниками птиц (см. ниже Филогенетическое положение).
Череп
В отличие от современных птиц, которые не имеют зубов, предчелюстные, челюстные и зубные кости археоптерикса несли зубы. Конические зубы археоптерикса в морфологии наиболее подобны зубам крокодилов.
Предчелюстные кости археоптерикса не срастались друг с другом, по контрасту с современными птицами. Верхняя и нижняя челюсть были лишены рогового чехла (рамфотеки), таким образом, археоптерикс не обладал клювом.
Большое затылочное отверстие, соединяющее полость черепа с позвоночным каналом, у археоптерикса находится сзади черепа, аналогично положению у дейнонихозавров, в то время как у современных птиц оно смещено на дно черепа.
Позвоночник
Шейные позвонки археоптерикса являются двояковогнутыми спереди и сзади, как у дейнонихозавров; в то время как у современных птиц шейные позвонки гетероцельные — имеют седловидные суставные поверхности.
Крестцовые позвонки археоптерикса не срастались друг с другом, тогда как у современных птиц поясничные, крестцовые и часть хвостовых позвонков срастаются в единую кость — сложный крестец. Крестцовый отдел позвоночника археоптерикса состоит из 5 позвонков, что сопоставимо числу крестцовых позвонков дейнонихозавров — 5—6. Крестцовый отдел позвоночника современных птиц включает 1 позвонок, входящий в состав сложного крестца.
21—23 несросшихся хвостовых позвонка археоптерикса образовывали длинный костистый хвост. У современных птиц, хвостовые позвонки срастаются в единую кость — пигостиль, служащую опорой для рулевых перьев.
Рёбра
Рёбра археоптерикса были лишены крючковидных отростков, которые у современных птиц обеспечивают прочность грудной клетки, скрепляя рёбра. По контрасту, в скелете археоптерикса присутствовали вентральные ребра (гастралии), типичные для многих пресмыкающихся, включая дейнонихозавров, но не встречающиеся у современных птиц.
Плечевой и тазовый пояса
Как у современных птиц и дейнонихозавров, ключицы археоптерикса срастались, образовывая вилочку.
Кости тазового пояса (подвздошная, седалищная и лобковая) у археоптерикса не срастались в единую кость, в отличие от состояния у современных птиц.
Лобковые кости археоптерикса незначительно обращены назад, как у дейнонихозавров, но не современных птиц. Также в отличие от современных птиц, лобковые кости археоптерикса заканчивались расширением в форме «ботинка», что характерно для дейнонихозавров. Кроме того, как и у дейнонихозавров, дистальные концы лобковых костей соединялись, образуя большой лобковый симфиз; у современных птиц лобковый симфиз отсутствует.
Передние конечности
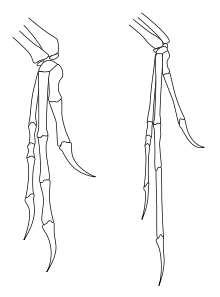 Кисти дейнониха (слева) и археоптерикса (справа).
Кисти дейнониха (слева) и археоптерикса (справа).Длинные передние конечности археоптерикса заканчивались тремя полноценно развитыми пальцами. Три пальца археоптерикса образованы двумя, тремя и четырьмя фалангами, соответственно. Та же фаланговая формула (2-3-4) типична для дейнонихозавров. По контрасту, трёхпалые пальцы современных птиц сильно редуцированы, число видоизмененных фаланг пальцев 1-2-1. Пальцы археоптерикса несли большие сильно изогнутые когти.
В запястье археоптерикса присутствовала полулунная кость, характерная для скелета современных птиц, известная также и у дейнонихозавров. Другие кости запястья и кости пясти не срастались в общую кость (пряжку), в отличие от современных птиц.
Задние конечности
Голень археоптерикса образована приблизительно равными в длине большой берцовой и малой берцовой костями, в то время как у современных птиц малая берцовая кость редуцирована и прирастает к большой берцовой. По контрасту с современными птицами, кости плюсны и часть костей предплюсны не срастались, таким образом, у археоптерикса не было цевки.
Основываясь на изучении лондонского и эйсштадского экземпляров, палеонтологи заключали, что большой палец задней конечности археоптерикса был полностью противопоставлен другим пальцам, что характерно для современных птиц. Однако анализ лучше сохраненного термополиского экземпляра показал, что в действительности ориентация большого пальца археоптерикса более подобна положению у дейнонихозавров, чем у современных птиц [3]. Впрочем, эта интерпретация была оспорена [5].
Кроме того, термополиский экземпляр подтвердил предположение о наличии у археоптерикса гиперудлиненного второго пальца задней конечности, несущего большой коготь [3]. Гиперудлиненный второй палец, снабженный большим хищным когтем, типичен для дейнонихозавров.
Оперение
 Первый рисунок берлинского экземпляра, сделанный неизвестным иллюстратором в 1878—1879 годах. Автор отметил присутствие в экземпляре отпечатков перьев.
Первый рисунок берлинского экземпляра, сделанный неизвестным иллюстратором в 1878—1879 годах. Автор отметил присутствие в экземпляре отпечатков перьев.Отпечатки перьев были ключевой особенностью, позволявшей классифицировать археоптерикса как птицу. Окаменелости птиц с отпечатками перьев редки. Сохранение перьев археоптерикса стало возможным благодаря литографическому известняку, в котором сохранились все экземпляры этого животного. Хотя отпечатки перьев присутствуют на всех экземплярах археоптерикса, сохранность у разных экземпляров перьев, как и костей, неодинакова. Наиболее информативны для изучения перьевого покрова археоптерикса лондонский и берлинский экземпляры [6]. Но поскольку подозревается существование нескольких видов археоптериксов, исследования перьев этих экземпляров не обязательно показательны для других видов.
Оперение археоптерикса в основных особенностях соответствует оперению современных и вымерших птиц. Археоптерикс обладал маховыми, рулевыми (функционально важными для полета) и контурными перьями, покрывавшими тело.
Маховые и рулевые перья археоптерикса образованы теми же структурными элементами, что и перья современных птиц (стержень пера, бородки первого порядка и бородки второго порядка, с отходящими от них крючочками) [6].
Маховые перья археоптерикса характеризуются асимметрией опахал, что типично для современных птиц, способных к полету. Рулевые перья были менее асимметричными. Отличие от современных птиц наблюдалось в отсутствии крылышка — отдельного подвижного пучка перьев на большом пальце передних конечностей.
Оперение тела археоптерикса менее хорошо описано, будучи должным образом исследованным только на хорошо сохранившемся берлинском экземпляре. Этот экземпляр на ногах носил «штаны» из хорошо развитых перьев, строение некоторых из них имело кое-какие отличия (например, отсутствовали бородочки, как у бескилевых птиц[6]), другие же были достаточно крепкими, допуская возможность полёта[7].
Вдоль спины был участок контурных перьев, симметричных и крепких (хотя и недостаточно жёстких по сравнению с полётными перьями), очень похожих на контурные перья на туловище современных птиц.
Остальные перья берлинского экземпляра имеют тип «псевдо-пуховых» и неотличимы от покровных волокон динозавра Sinosauropteryx: мягкие, разрозненные и, возможно, даже внешне более похожие на мех — они покрывали все оставшиеся части туловища (там, где сохранились), а также нижнюю часть шеи[6].
Нет никаких признаков оперения на верхней части шеи и голове. Хотя они могли и отсутствовать, как у многих оперённых динозавров, это может быть и недостатком сохранности экземпляров: похоже, что большинство экземпляров археоптерикса попало в осадочные породы после некоторого времени пребывания на поверхности моря, плавая на спине. Голова, шея и хвост обычно изогнуты книзу, что предполагает, что экземпляры только начинали разлагаться, когда были погребены. Сухожилия и мускулы расслабились и тела приняли характерную для обнаруженных экземпляров форму. Также это означало бы, что кожа к тому моменту размягчилась и обвисла. Это предположение поддерживается фактом, что у некоторых экземпляров полётные перья начинали отваливаться к моменту погружения в осадочную породу. Таким образом, перья головы и шеи могли просто отпасть, тогда как более крепко держащиеся хвостовые перья остались[8].
Мозг и внутреннее ухо
Отличительной особенностью черепов птерозавров, некоторых тероподов и птиц являются тонкие мозговые оболочки и маленькие венозные пазухи, что делает возможной точную оценку поверхностной морфологии, объёма и массы мозга вымерших представителей этих таксонов. Лучшая на сегодняшний день реконструкция мозга археоптерикса была сделана в 2004 году в Университете Техаса с использованием рентгеновской томографии [9].
Объём мозга археоптерикса приблизительно в 3 раза больше, чем у рептилий сходного размера и в сравнительном отношении соответствует объёму мозга современных птиц. По контрасту, полушария мозга пропорционально меньше, чем у современных птиц, и не окружают обонятельные тракты.
Форма зрительных долей мозга типична для современных птиц, но сами зрительные доли расположены более фронтально. Мозжечок археоптерикса в сравнительном отношении больше, чем у родственных ему тероподов, но меньше, чем у современных птиц.
Передний и боковой полукружные каналы находятся в типичном для архозавров (а не для птиц) положении. Однако, как и у птиц, передний полукружный канал значительно удлинен и загнут в обратном направлении.
Таким образом, в строении мозга археоптерикса прослеживается наличие как рептилийных, так и птичьих особенностей. Увеличение зрительных долей и мозжечка является адаптацией, без которой невозможен полет [9].
Палеоэкология
 Современный саговник.
Современный саговник.В позднем юрском периоде территория современной Европы была архипелагом островов в мелководном теплом тропическом море, располагаясь гораздо ближе к экватору, чем в настоящее время. Зольнхофенские известняки, которым принадлежат все обнаруженные экземпляры археоптерикса, позволяют реконструировать картину эпохи позднего юрского периода. Следы флоры, хотя и довольно скудные, включают саговники и хвойные растения. Останков животного мира больше, обнаружено множество насекомых, маленькие ящерицы, птерозавры и компсогнаты[10].
Превосходная сохранность останков археоптериксов и других наземных животных, обнаруженных в зольнхофенских известняках, свидетельствует, что останки не могли быть перенесены издалека к месту фоссилизации[11]. То есть, обнаруженные экземпляры археоптериксов скорее обитали на этих низких островах, окружающих зольнхофенскую лагуну, а не были занесены сюда после смерти откуда-то из другого места.
Популяции археоптерикса отличались малочисленностью. В отложениях у Зольнхофена скелеты археоптериксов встречаются гораздо реже останков рамфоринхов, представителей группы птерозавров, доминировавших в экологической нише, в настоящее время занятой морскими птицами[12].
На невысоких островах, окружавших зольнхофенскую лагуну, преобладал полупустынный субтропический климат с длинным засушливым периодом и незначительными осадками. Приспособленная к такому климату флора состояла преимущественно из невысоких (до 3 метров) кустарников[12]. В противоположность распространённой реконструкции археоптерикса, взбирающегося на большое дерево, на островах, видимо, почти отсутствовали высокие деревья. В отложениях обнаружено очень мало древесных стволов и не найдено окаменевшей пыльцы деревьев.
Трудно реконструировать образ жизни археоптериксов. Относительно этого существует несколько теорий. Некоторыми исследователями было высказано предположение, что археоптериксы в основном были адаптированы к наземному образу жизни[13], тогда как другие предполагают, что образ жизни археоптериксов главным образом, древесный. Отсутствие деревьев не противоречит этому предположению — некоторые современные виды птиц живут исключительно в невысоких кустарниках. Разные аспекты морфологии археоптерикса указывают как на наземное, так и на древесное существование. Длина ног и удлинённые стопы позволили некоторым авторам прийти к выводу об универсальности археоптерикса, который мог кормиться как в зарослях, так и на земле и даже вдоль побережья лагуны[12]. Вероятнее всего добыча была небольшой, совсем маленьких жертв археоптерикс хватал челюстями, тех, кто побольше — когтями.
На основании анализа строения глазниц высказывается предположение, что археоптерикс вёл ночной образ жизни[14].
Способность к полёту
Асимметрия маховых перьев свидетельствует, что археоптерикс был аэродинамически адаптирован к полёту. Но археоптерикс не обладал рядом других адаптивных особенностей, характерных для способных к полету птиц, современных и вымерших, поэтому механика его полета и сам полет были более примитивными, по сравнению с более поздними птицами [15].
Среди исследователей нет единого мнения по вопросу о том, был ли археоптерикс способен к машущему (активному) полету или только к планирующему (пассивному).
Отсутствие у археоптерикса киля, надкоракоидного сухожилия, а также малый угол плечевого сустава и оцененная нагрузка на крыло, по мнению некоторых исследователей, свидетельствуют, что археоптерикс был способен только к планирующему полету. Боковая ориентация сустава между лопаткой, коракоидом и плечевой костью показывает, что археоптерикс не способен был поднимать крылья выше уровня спины — необходимое условие для замаха крыла у современных птиц [15]. Была выдвинута гипотеза, что его планирующий полёт сопровождался небольшими движениями крыльев без замаха[16].
Другие исследователи отмечают, что археоптерикс отличается от типичных преимущественно планирующих птиц в форме тела, а также в размере крыльев. Кроме того, они указывают, что костная грудина или бумерангообразная вилочка, или пластинообразный коракоид археоптерикса могли служить местом прикрепления мышц, двигавших крыло. Сторонники таких аргументов заключают, что археоптерикс был способен к некоторой форме примитивного машущего полета [15].
Поскольку реконструкция функциональной анатомии вымершего животного, находящегося на промежуточной эволюционной стадии развития, проблематична, в дискуссии о способности археоптерикса к полету вряд ли в скором будущем возобладает та или иная точка зрения.
Таксономия
Исторически первым названием археоптерикса является Pterodactylus crassipes Ван Мейера. Название Archaeopteryx lithographica изначально было дано единственному перу, описанному Ван Мейером. В 1960 Суинтон предложил, чтобы название A. lithographica было официально отнесено к лондонскому экземпляру [17]. Международная комиссия по зоологической номенклатуре устранила множество альтернативных названий в пользу A. lithographica [18].
Единой таксономии археоптериксов нет. Вопрос о том, к скольким видам и родам принадлежат известные экземпляры, остается открытым. Большинство экземпляров классифицировалось как представители отдельных видов или родов. Берлинский экземпляр классифицировался как Archaeornis siemensii, айхштетский — как Jurapteryx recurva, мюнхинский — A. bavarica, зольнхофенский — Wellnhoferia grandis.
Экземпляры отличаются друг от друга в ряде существенных характеристик (больший или меньший размер тела, различные пропорции пальцев, более (или менее) суженные морды и т. д.). Такие различия могут быть интерпретированы как особенности, отличающие один вид (род) от других. Однако они также могут быть приписаны половому диморфизму или разным возрастным стадиям развития животных. Из всех предложенных вариантов, наибольшее признание получили A. lithographica, A. siemensii, A. bavarica, Wellnhoferia grandis. В 2007 термополиский экземпляр был классифицирован как A. siemensii [4].
В 1867 Томас Генри Хаксли поместил археоптерикса в подкласс Sauriurae (ящерохвостые птицы) [19]. Эта классификация принималась на протяжении долгого времени. В конце XX века к Sauriurae также отнесли энанциорнисов (Enantiornithes) и конфуциусорнисов (Confuciusornithidae). Однако монофилия Sauriurae была подвергнута сомнению, и некоторые современные исследователи не признают этот таксон. Они отнесли археоптерикса к семейству Archaeopterygidae в отряде Archaeopterygiformes.
Синонимы
 Некоторые виды археоптериксов, рядом с ногами человека для сравнения.
Некоторые виды археоптериксов, рядом с ногами человека для сравнения.- Pterodactylus crassipes Meyer, 1857
- Rhamphorhynchus crassipes (Meyer, 1857) (as Pterodactylus (Rhamphorhynchus) crassipes)
- Archaeopteryx lithographica Meyer, 1861 [nomen conservandum]
- Scaphognathus crassipes (Meyer, 1857) Wagner, 1861
- Griphosaurus problematicus Wagner, 1861 [nomen oblitum 1961 per ICZN Opinion 607]
- Griphornis longicaudatus Woodward, 1862 [nomen oblitum 1961 per ICZN Opinion 607]
- Griphosaurus longicaudatus (Owen, 1862) [nomen oblitum 1961 per ICZN Opinion 607]
- Archaeopteryx macrura Owen, 1862 [nomen oblitum 1961 per ICZN Opinion 607]
- Archeopteryx macrurus Owen, 1863
- Archaeopteryx siemensii Dames, 1897
- Archaeornis siemensii (Dames, 1897) Petronievics, 1917
- Archaeopteryx oweni Petronievics, 1917 [nomen oblitum 1961 per ICZN Opinion 607]
- Archaeopteryx crassipes (Meyer, 1857) Ostrom, 1972
- Archaeopteryx recurva Howgate, 1984
- Jurapteryx recurva (Howgate, 1984) Howgate, 1985
- Archaeopteryx bavarica Wellnhofer, 1993
- Wellnhoferia grandis Elżanowski, 2001
Филогенетическое положение
Поскольку археоптерикс был и остается наиболее ранней и наиболее примитивной известной птицей, вопрос о его филогенетическом положении исторически был и остается неразрывно связанным с изучением происхождения птиц. На основе исследований экземпляров археоптерикса выдвигались различные гипотезы филогенетических отношений птиц, но общепринятой филогении нет.
- Текодонтная гипотеза
Эту гипотезу предложил в начале XX века Герхард Хейльман (Gerhard Heilmann), основываясь на детальном обзоре биологии птиц (включая археоптерикса) и рептилий. По его мнению, наиболее близкими родственниками птиц следует считать триасовых псевдозухий (из надотряда текодонтов). Текодонтная гипотеза была общепринятой в середине XX столетия и поддерживается сегодня рядом специалистов. Однако вплоть до настоящего времени среди псевдозухий не удалось обнаружить форм филогенетически близких к археоптериксу или другим ранним птицам.
- Гипотеза Уокера
А Уокер обосновывал в 70-х годах XX века родство археоптерикса и всех птиц с триасовыми архозаврами из подотряда сфенозухий, родственных современным крокодилам. Аргументы Уокера неоднократно критиковались, и сам автор впоследствии признал гипотезу несостоятельной.
- Тероподная гипотеза
Гипотеза первоначально была выдвинута Томасом Генри Хаксли в XIX веке. В 70-ых годах XX века её возродил Джон Остром. Основываясь на сравнении пересмотренной им остеологии археоптерикса и тероподных динозавров (в особенности описанного им дейнониха), Остром заключил, что ближайшими родственниками археоптерикса являются целурозавры из семейства дромеозаврид.
Впоследствии выводы Острома были подтверждены кладистическими исследованиями, обнаружением экземпляров целурозавров с отпечатками перьев, сравнением микроструктуры яичной скорлупы и репродуктивной биологии целурозавров и птиц [15][20][21]. Сегодня тероподная гипотеза принимается большинством палеонтологов.
По мнению сторонников тероподной гипотезы, о родстве археоптерикса с тероподами свидетельствует целый ряд продвинутых признаков (синапоморфий)[13][20]:
- расширенность слезной кости в дорзальной части;
- краниально направленный медиальный мыщелок нижнечелюстного отдела квадратной кости;
- большой эктоптеригоид;
- одинаковая ширина шейки и тела лопатки по всей длине;
- плоский субквадратный коракоид;
- заметный бугор на коракоиде кранио-вентрально от гленоидного отверстия;
- перпендикулярное положение плоскости проксимального эпифиза плеча к плоскости дистального эпифиза плеча;
- короткое предплечье (короче кисти и плечевой кости);
- сохранение в кисти 1-3-го пальцев и редукция 4-5-го;
- контакт полулунной карпалии с 1 и 2 метакарпалиями;
- увеличенные когтевые фаланги кисти с большими буграми для прикрепления сухожилий сгибателей;
- развитие малого трохантера в проксимальной части бедра;
- краниальная часть подвздошной кости заметно больше каудальной;
- развитие ножки на подвздошной кости для сочленения с лобковой костью;
- вильчатый каудальный конец подвздошной кости;
- большой лобковый симфиз.
Современная филогения первых птиц и наиболее близких к ним групп тероподов выглядит следующим образом:
Coelurosauria Целурозавры Tetanurae Maniraptoriformes Maniraptora Eumaniraptora Deinonychosauria (Dromaeosauridae + Troodontidae) Aves Птицы Archaeopterygiformes Pygostylia
Значение
Лондонский экземпляр археоптерикса был обнаружен в 1861 году, два года спустя после опубликования «Происхождения видов» Чарльза Дарвина. Реконструкция анатомии археоптерикса, основанная на лондонском экземпляре, показала, что по строению скелета он занимает промежуточное положение между рептилиями и птицами. В «Происхождении видов» Дарвин отмечал, что, согласно его теории, такие формы должны были существовать, и указывал на их отсутствие как на серьёзное возражение против теории. В связи с этим открытие археоптерикса привлекло внимание единомышленников Дарвина (в первую очередь Томаса Генри Хаксли). Сам Дарвин в переизданиях «Происхождения видов» кратко упоминал археоптерикса в десятой главе, посвященной неполноте палеонтологической летописи (см. например, издание 1866 года [22].). В итоге археоптерикс и его анатомия стали предметом дискуссий между сторонниками эволюционного учения и их оппонентами. Открытие археоптерикса на заре становления эволюционной биологии и реконструкция его скелета сделали археоптерикса хрестоматийным примером существа, анатомия которого позволяет проследить происхождение одного таксона от другого. На сегодняшний день археоптерикс является одним из наиболее известных за пределами научного сообщества вымерших животных.
Археоптерикс был и остается самой ранней и самой примитивной птицей из известных науке. Поэтому любое исследование анатомии и физиологии ранних птиц, их происхождения, эволюции, филогении, возникновения полета, основывались и основываются в первую очередь на экземплярах археоптерикса. В связи с этим биология, в особенности анатомия, археоптерикса многократно переоценивалась и пересматривалась палеонтологами нескольких поколений. На посвященной археоптериксу конференции, состоявшейся в 1984, было отмечено, что за всю историю палеонтологии не многие вымершие животные становились предметом специальной конференции.
Для сохранения в окаменелостях покровных тканей необходимы особые условия, и поэтому окаменелости животных со следами наружного покрова редки. Экземпляры археоптерикса сохраняют отпечатки перьев, наличие которых стало основной диагностической особенностью, позволившей классифицировать археоптерикса как птицу. Впоследствии исследования экземпляров археоптерикса позволили реконструировать оперение этой птицы и выдвигать гипотезы об эволюции перьев.
Подлинность
В 80-х годах XX века подлинность археоптерикса была поставлена под сомнение рядом авторов, включая известного британского астронома Фреда Хойла и американского физика Ли Спетнера (Lee Spetner). Начиная с 1985 года эти авторы опубликовали ряд статей, в которых утверждали, что перья берлинского и лондонского экземпляров археоптерикса были фальсифицированы путём добавления к подлинным окаменелостям перьев современных птиц [23][24][25][26]. Согласно Хойлу с соавторами без добавленных перьев экземпляры археоптерикса классифицировались бы как останки динозавра компсогнатуса (единственный известный в то время динозавр в золонгофенских отложениях).
Их аргументы были опровергнуты ведущим палеонтологом Лондонского музея естествознания Аланом Дж. Чериджем (Alan J. Charig; 1927—1997) и соавторами[27]. Большинство из доказательств фальсификации основывалось на неосведомлённости о процессах литификации. Высказывались сомнения в том, что пласты могут разделяться так гладко или что содержащая окаменелости половина пласта могла так хорошо сохраниться, тогда как вторая половина — нет[23][25]. Однако, это обычные свойства золонгофенских ископаемых, мертвые животные падали на затвердевшую поверхность, которая формировала естественную плоскость для следующих пластов, так что большая часть останков оставалась на одной стороне, и совсем мало впечатывалось в твердую поверхность другой[27]. Также они неправильно интерпретировали окаменелости[24], ошибочно утверждали, что другие экземпляры, известные на то время не имели перьев[23][24]. Однако максбергский и эйнштадтский экземпляры имеют явные отпечатки перьев[27]. Кроме того, новые экземпляры, обнаруженные уже после заявления о подделке, также сохранены с отпечатками перьев. Экземпляры археоптерикса многократно исследовались множеством профессиональных палеонтологов, и никто из них не классифицировал какой-либо экземпляр как компсогнатуса (см. раздел таксономия), равно как никто из них не утверждал, что экземпляры были подделаны.
Черидж и соавторы указали на микротрещины на литографических плитах, бегущие через обе стороны плиты и ископаемые отпечатки, которые существовали ещё до открытия останков, как свидетельство того, что перья были с самого начала[27]. В ответ на это Ли Спетнер и соавторы попытались показать, что трещинки могли бы естественным образом распространиться на предполагаемый слой цемента[28], но пренебрегли фактом, что трещинки были старыми, давно заполнились кальцитом и не могли распространяться[29]. Они также попытались показать присутствие цемента на лондонском экземпляре при помощи рентгенографии и действительно обнаружили нечто, что не являлось камнем[28]. Однако, это было и не цементом, скорее всего являясь фрагментом кремниевого мусора, оставшегося после того, как был сделан слепок[29].
Наконец, предполагаемые ими мотивы для фальсификации были слабыми и противоречивыми. Одним из мотивов они называли стремление Ричарда Оуэна поддержать теорию Дарвина, что маловероятно, учитывая взгляды Оуэна на Дарвина и его теорию. Другой мотив — это желание Оуэна установить ловушку для Дарвина, в надежде, что последний поддержит окаменелости, а Оуэн сможет тогда его дискредитировать подделкой. Это также маловероятно, так как Оуэн сам написал детальный труд по лондонскому экземпляру, и подобная дискредитация обернулась бы против него самого[29].
Доводы сторонников фальсификации не были всерьёз восприняты палеонтологами, так как их доказательства основывались на непонимании геологии, и никогда не затрагивали другие ископаемые с перьями, число находок которых значительно увеличилось с того времени.
См. также
Примечания
- ↑ В 1984 году палеонтолог Шанкр Чаттерджи обнаружил ископаемые останки возрастом 225—210 млн лет, которые, по его мнению, являются останками птицы, филогенетически более близкой к современным птицам, чем археоптерикс. Род позвоночных получил название протоавис. Однако его отношение к птицам подвергнута сомнению.
- ↑ Mäuser M. Der achte Archaeopteryx. Fossilien, 1997, v. 3, p. 156—157
- ↑ 1 2 3 Mayr G, Pohl B & Peters DS. A well-preserved Archaeopteryx specimen with theropod features. Science, 2005, v. 310 p. 1483—1486
- ↑ 1 2 Mayr G. et al. The tenth specimen of Archaeopteryx. Zoological Journal of the Linnean Society, 2007, v. 149, p. 97-116
- ↑ Feduccia A. et al. Archaeopteryx 2007: Quo Vadis? The Auk, v. 124, p. 373—380
- ↑ 1 2 3 4 Christensen P, Bonde N. (2004). Body plumage in Archaeopteryx: a review, and new evidence from the Berlin specimen. Comptes Rendus Palevol. 3: 99-118. PDF fulltext
- ↑ Longrich N. (2006): Structure and function of hindlimb feathers in Archaeopteryx lithographica. Paleobiology. 32(3): 417—431. DOI:10.1666/04014.1 (HTML abstract)
- ↑ Elżanowski A. (2002): Archaeopterygidae (Upper Jurassic of Germany). In: Chiappe, L. M. & Witmer, L. M (eds.), Mesozoic Birds: Above the Heads of Dinosaurs: 129—159. University of California Press, Berkeley.
- ↑ 1 2 P. D. Alonso et al. The avian nature of the brain and inner ear of Archaeopteryx. Nature, 2004, v. 430, p. 666—669
- ↑ Chiappe, Luis M. (2007). Glorified Dinosaurs. Sydney: UNSW Press, 118—146. ISBN 0-471-24723-5.
- ↑ Davis, P.; and Briggs, D. (1998). «Влияние разложения и дезартикуляции на сохранность ископаемых птиц/The impact of decay and disarticulation on the preservation of fossil birds». Palaios 13 (1): 3-13. (англ.)
- ↑ 1 2 3 Paul, Gregory S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. ISBN 0-8018-6763-0.
- ↑ 1 2 Ostrom, J.H. (1976). «Archaeopteryx and the origin of birds». Biological Journal of the Linnean Society 8: 91-182.
- ↑ Archaeopteryx may have hunted at night. New Scientist.
- ↑ 1 2 3 4 Padian K., Chiappe L. The origin and early evolution of birds. Biol. Rev., 1998, v. 73, p. 1-42
- ↑ Senter, P. (2006). Scapular orientation in theropods and basal birds and the origin of flapping flight. Acta Palaeontologica Polonica. 51(2): 305—313. PDF fulltext
- ↑ Swinton, W. E. Opinion 1084, Proposed addition of the generic name Archaeopteryx VON MEYER, 1861 and the specific name Lithographica, VON MEYER, 1861, as published in the binomen Archaeopteryx Lithographica to the official lists (Class Aves). Bulletin of Zoological Nomenclature, 1960, v. 17(6-8) p. 224—226.
- ↑ ICZN. Opinion 607, Archaeopteryx VON MEYER, 1861 (Aves); Addition to the Official list. Bulletin of Zoological Nomenclature, 1961, v. 18(4), p. 260—261
- ↑ Huxley, T. H. On the classification of birds and on the taxonomic value of the modifications of certain of the cranial bones observable in that class. Proceedings of the Zoological Society of London, 1867, p. 415—472.
- ↑ 1 2 Gauthier J. «Saurischian Monophyly and the origin of birds», in K. Padian: The Origin of Birds and the Evolution of Flight. 1986. Mem. California Acad. Sci 8, 1-55
- ↑ Chiappe L. The Closest Relatives of Birds. Ornitología Neotropical, 2004, v. 15, p. 101—116
- ↑ Darwin C. R. On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. London: John Murray 1866. Четвертое издание
- ↑ 1 2 3 Hoyle, F.; Wickramasinghe, N.C.; and Watkins, R.S. (1985). «Archaeopteryx». British Journal of Photography 132: 693–694.
- ↑ 1 2 3 Watkins, R.S.; Hoyle, F.; Wickrmasinghe, N.C.; Watkins, J.; Rabilizirov, R.; and Spetner, L.M. (1985). «Archaeopteryx - a photographic study». British Journal of Photography 132: 264–266.
- ↑ 1 2 Watkins, R.S.; Hoyle, F.; Wickrmasinghe, N.C.; Watkins, J.; Rabilizirov, R.; and Spetner, L.M. (1985). «Archaeopteryx - a further comment». British Journal of Photography 132: 358–359, 367.
- ↑ Watkins, R.S.; Hoyle, F.; Wickrmasinghe, N.C.; Watkins, J.; Rabilizirov, R.; and Spetner, L.M. (1985). «Archaeopteryx - more evidence». British Journal of Photography 132: 468–470.
- ↑ 1 2 3 4 Charig, A.J.; Greenaway, F.; Milner, A.N.; Walker, C.A.; and Whybrow, P.J. (1986). «Archaeopteryx is not a forgery». Science 232: 622–626.
- ↑ 1 2 Spetner, L.M.; Hoyle, F.; Wickramasinghe, N.C.; and Magaritz, M. (1988). «Archaeopteryx - more evidence for a forgery». The British Journal of Photography 135: 14–17.
- ↑ 1 2 3 Nedin, Chris On Archaeopteryx, Astronomers, and Forgery (15 декабря 2007). Архивировано из первоисточника 8 февраля 2012. Проверено 17 марта 2007.
Рекомендуемая литература
- Кэрролл Р. Палеонтология и эволюция позвоночных. Т. 3. — Пер. с англ. — М.: Мир, 1993.
- Е. Н. Курочкин. Параллельная эволюция тероподных динозавров и птиц. Зоологический журнал, 2006, т. 85 (№ 3), с. 283—297
- Ostrom J. Archaeopteryx. Discovery, 1975, V. 11, p. 15-23.
- de Beer, G.R. (1954). Archaeopteryx lithographica: a study based upon the British Museum specimen. Trustees of the British Museum, London.
- Chambers, P. (2002). Bones of Contention: The Fossil that Shook Science. John Murray, London. ISBN 0-7195-6059-4.
- Feduccia, A. (1996). The Origin and Evolution of Birds. Yale University Press, New Haven. ISBN 0-300-06460-8.
- Heilmann, G. (1926). The Origin of Birds. Witherby, London.
Ссылки

Археоптерикс на Викискладе? - Археоптерикс // Энциклопедический словарь Брокгауза и Ефрона: В 86 томах (82 т. и 4 доп.). — СПб., 1890—1907.
- А. В. Марков. Доклад, прочитанный в институте Общей Генетики 18 марта 2004 г.
- Е. Н. Курочкин. Новый взгляд на эволюцию птиц
- Доказано, что археоптерикс летал
- Фотографии экземпляров археоптерикса
- Кэрролл Р. Палеонтология и эволюция позвоночных. — Пер. с англ. — М.: Мир, 1993
- Могла ли первоптица летать? на Dino-Millennium

Категории:- Животные по алфавиту
- Фауна юрского периода
- Животные, описанные в 1861 году
- Переходные формы
- Вымершие животные







Wikimedia Foundation. 2010.